Бюргерс векторы,оның шамасы мен бағыты.Дислокация қозғалысы.
ВЕКТОРЫ
Векторы
Векторы
Векторы в пространстве
Векторы и линейные операции над ними
Векторы и операции над ними.
Векторына салынған параллелепипедтің көлемін табыңыз
Векторының бірлік векторын табыңыз
Векторы на основе Ti-плазмид. Некоторые виды агробактерий (Agrobacteria) могут заражать двудольные растения, вызывая образование опухолей — корончатых галлов. Одним из самых сильных индукторов опухолей служит почвенная бактерия A. tumefaciens. Способность этой бактерии к образованию опухоли связана с большой внехромосомной плазмидой, получившей название Ti-плаз- мида (от ангд. tumor inducing— индуцирующие опухоль). Ti-плас миды — это естественные векторы для генов, обладающие всем* функциями, необходимыми для переноса, стабильного в ключе ния и экспрессии генетической информации в растениях. Ош_ имеют широкий круг хозяев. В бактериальных клетках Ti-плазми- ды реплицируются автономно. Эти плазмиды различаются по тигг кодируемых опинов — белков, которые используются бактериями в качестве источников азота и углерода. Обычно встречаются плазмиды, кодирующие два типа опинов: либо октопин (октопи новая плазмида), либо нопалин (нопалиновая плазмида).
Промежуточный и бинарный векторы. Эти векторы конструируются на основе Ti-плазмид. Промежуточный вектор получают путем ряда сложных операций. Сначала Т-область с помощью рестриктаз вырезают из плазмиды, вставляют в вектор для клонирования в клетке Е. coli и размножают. Затем внутрь Т-области встраивают чужеродный ген и вновь размножают. Полученную реком- бинантную плазмиду вводят в клетки A. tumefaciens, несущие полную Ti-плазмиду. В результате двойного кроссинговера между гомологичными участками Т-область рекомбинантной плазмиды, содержащая чужеродный ген, включается в Ti-плазмиду клетки хозяина, заместив в ней нормальную Т-область. Наконец, бактериями, имеющими Ti-плазмиду со встроенными генами, заражают растения, где эти гены встраиваются в геном растительной клетки.
Бинарные векторы представляют собой бактерии, содержащие две разные Ti-плазмиды. Одна из них несет vir-область и обеспечивает интеграцию в геном растительной клетки Т-области, содержащей любые гены другой плазмиды. В этом случае двойной кроссинговер не требуется.
Однако полученные в результате заражения бактериями растительные клетки не способны к регенерации, так как у них подавлена дифференцировка. Трансформированные растительные клетки смогут дифференцироваться, если в гены, блокирующие диффе- ренцировку, ввести мутации или вырезать их из Т-ДНК.
Векторы на основе ДНК-содержащих вирусов растений. Вирусы можно рассматривать как разновидности чужеродной нуклеиновой кислоты, которые реплицируются и экспрессируются в клетках растений. Подавляющее большинство фитовирусов в качестве носителя генетической информации содержат РНК. Только 1 — 2 % чирусов, инфицирующих растения, относятся к ДНК-содержа- щим. Именно эти вирусы удобны для использования в технологии Рекомбинантных ДНК, а также в качестве векторов.
ДНК-содержащие вирусы могут включать одноцепочечную или двухцепочечную ДНК. В качестве представителей первой группы можно назвать вирус золотистой мозаики фасоли (ВЗМФ| или вирус полосатости кукурузы. Наиболее изученный представитель группы вирусов с двухцепечечной ДНК — вирус мозаики цветной капусты (ВМЦК), поражающий в основном растения’ семейства крестоцветные.
Обычно фитовирусы реплицируются с образованием большое го числа копий молекул нуклеиновых кислот — 106 и более молекул на зараженную клетку. Поэтому фитовирусы представляют собой очень эффективные средства для получения хорошей экспрессии чужеродного гена. Кроме высокой копийности вирусной нуклеиновой кислоты вирусные векторные системы имеют еще ряд преимуществ: малый размер генома (возможность легкой манипуляций вирусной ДНК) и сильные промоторы, обеспечивающие эффективную экспрессию чужеродных генов.
Однако вирусы в качестве векторов обладают и существенным^ недостатками: имеют небольшую емкость, патогенны и неспособ ны встраиваться в хромосомы хозяина. Небольшую емкость можно увеличить, если инфицировать вирусом (например, ВМЦК растительные протопласты, а не клетки. В этом случае инфекция не передается от клетки к клетке, нет необходимости в упаковке ДНК в вирусные частицы.
Следовательно, часть вирусного генома, ответственная за упаковку в вирусные частицы, может быть удалена и замещена до, полнительной чужеродной ДНК. Другой недостаток — отсутствий способности встраиваться в геном растительной клетки — удается обойти (по крайней мере для ВМЦК) благодаря специальном® методическому приему — агроинфекции. Для этого геном ВМЦ1 встраивают в Т-область Ti-плазмиды и в ее составе интегрируют ядерный геном различных растений.
141. Организация и «поведение» Ti- плазмиды.
Некоторые виды агробактерий (Agrobacteria) могут заражать двудольные растения, вызывая образование опухолей — корончатых галлов. Одним из самых сильных индукторов опухолей служит почвенная бактерия A. tumefaciens. Способность этой бактерии к образованию опухоли связана с большой внехромосомной плазмидой, получившей название Ti-плаз- мида (от ангд. tumor inducing— индуцирующие опухоль). Ti-плас миды — это естественные векторы для генов, обладающие всем* функциями, необходимыми для переноса, стабильного в ключе ния и экспрессии генетической информации в растениях. Ош_ имеют широкий круг хозяев. В бактериальных клетках Ti-плазми- ды реплицируются автономно. Эти плазмиды различаются по тигг кодируемых опинов — белков, которые используются бактериями в качестве источников азота и углерода. Обычно встречаются плазмиды, кодирующие два типа опинов: либо октопин (октопи новая плазмида), либо нопалин (нопалиновая плазмида).
После заражения часть Ti-плазмиды встречается в хромосома; клеток растения-хозяина. Следовательно, A. tumefaciens встраивает часть своего генома в ДНК растительной клетки и заставляет et- таким способом изменять метаболизм, синтезируя вещества, не обходимые для бактерий. Именно это свойство A. tumefaciens i послужило поводом для создания на основе Ti-плазмиды вектора, доставляющего необходимые гены в клетку.
Участок Ti-плазмиды, встречающийся в хромосомах растительных клеток, называется Т-областью в бактерии и Т-ДНК в клетках растений. Т-область включает примерно 10 % Ti-плазмиды и содержит гены, отвечающие за индукцию опухоли, синтез опинов и подавление дифференцировки (гормоннезависимый рост клеток). Важно отметить, что все гены, ответственные за перенос и интеграцию генов Т-области, находятся не в ней самой, а рядом — в области вирулентности — vir-области (рис. 5.17).
Т-области ограничены прямыми повторяющимися последовательностями, и любая ДНК, вставленная между этими повторами, будет принята за Т-область и перенесена в растительную клетку Недостаток этих плазмид состоит в том, что некоторые гены находящиеся в Т-ДНК, заставляют расти клетки растений незави симо от гормонов, вносимых в питательную среду, на которой куль
тивируются данные клетки. В свя зи с этим очень трудно регене рировать нормальное растение и клеток, содержащих полнук, последовательность Т-ДНК. Др> гой недостаток — большие par меры
С помощью таких векторов наблюдают временную экспрессию клонир. геновTi-плазмиды, из-за коте рых затруднены какие-либо м. нипуляции с ней, поэтому вст вить ген в плазмиду традицио! ными методами невозможно.
В настоящее время констр ируются производные Ti-пла миды, в которых оставляют р гуляторный участок Т-обласг а вместо ее структурных генов вшивают структурную часть гена, который надо ввести в растение. Такие гены с позиции их регенерации безвредны для растений.
Существуют и другие бактерии (A. rhizogenes), вызывающие усиленное образование корешков при заражении растений. За этот процесс ответственны содержащиеся в них так называемые Ri- плазмиды (от англ. root inducing — индуцирующий корни). Ri-плаз- миды выгодно отличаются от Ti-плазмид тем, что они служат естественными безвредными векторами, так как трансформированные с их помощью растительные клетки сохраняют способность к морфогенезу и к регенерации здоровых растений. В связи с этим Ri-плазмиды в данный момент рассматриваются как более перспективные векторы.
Дата добавления: 2015-04-16 ; просмотров: 108 ; Нарушение авторских прав
Наиболее пригодными яаляется Caulimoviridae — каулимовирусы.(их генома, представляет собой двунитевую ДНК до 8300 п.н.). Их ДНК имеет вид кольцевой ковалентко замкнутой суперскрученной молекулы. Эта ДНК транскрипционно активна и основными транскриптами являются 35S и 19S РНК. 35S РНК соответствует по длине всему геному вируса и служит матрицей для синтеза ДНК, которая может упаковываться в вирусные капсиды. Наличие в геноме вирусов гена обратной транскриптазы и этапа обратной транскрипции в жизненном цикле сближает каулимовирусы с ретровирусами животных, из-за чего их объединяют в супергруппу параретровирусов.
Сейчас известно более десятка каулимовирусов. Наиболее изученным вирус мозаики цветной капусты.
Успешными примерами экспрессии чужеродных для растений генов с вирусных векторов на основе каулимовирусов были:
С помощью таких векторов можно вводить в растения только небольшие фрагменты (не более 300 п.н.), и что полученные путем вирусной трансформации растения не способны передавать приобретенный признак в рядах поколений.
Можно прочитать еще про вирус мозаики цветной капусты..минимально..это билет 9, 1 вопрос.
Билет 9
Генет.инженерия вируса мозаики цветной капусты
Наиболее изученным вирусом является вирус мозаики цветной капусты.В вирусных частицах ДНК CaMV представляет собой кольцевую, но не ковалентно замкнутую молекулу.
В ее составе имеются 7 открытых рамок считывания. Рамка I несет информацию о белке, обеспечивающем перенос вирусных частиц по плазмадесмам из одной растительной клетки в другую, то есть системное распространение по растению. Рамки II и III кодируют протеины Р2 и Р3, которые отвечают за перенос вирионов тлями от растения к растению и называются факторами трансмиссии. Рамка IV соответствует предшественнику капсидных белков, рамка V – предшественнику протеиназы, обратной транскриптазы и РНКазы Н, которые нужны для построения ДНК на матрице 35S РНК. Рамка VI кодирует белок телец включения, которые появляются в цитоплазме инфицированной растительной клетки и служат местом синтеза геномной ДНК и сборки вирусных частиц. Для белка, кодируемого рамкой VII функции не известна.С точки зрения генетической инженерии особый интерес представляют две межгеномные области , в которых находятся промоторы 19S РНК и 35S РНК. Именно в эти области осуществляли вставки фрагментов ДНК с целью оценки возможностей использования каулимовирусов как векторных молекул для генетической трансформации растений. Помимо межгеномных областей пригодными для инсерций генетического материала без последующего нарушения цикла репродукции вируса являются рамки считывания II и III.
Получение и применение трансгенных овец
Нужно для использования их молока для получения медицинских препаратов, поскольку генетические конструкции для экспрессии генов человека в клетках молочной железы этих животных получены и апробированы. В гибридных ДНК используются промоторы гена бетта-лактоглобулина, гена бетта-казеина и гена альфа S1-казеина, а среди клонированных генов человека – ген активатора плазминогена, ген альфа-антитрипсина (при отсутствии этого белка развивается эмфизема, дефицит ингибитора сывороточных протеаз, поражение легких, цирроз печени), ген фактора IX системы свертывания крови (белок нужен для лечения гемофилии В), ген лактоферрина, ген интерлейкина-2. Из трансгенного мелкого рогатого скота упоминают только овец с повышенной скоростью роста шерсти. Этот вариант был получен путем введения кДНК гена инсулиноподобного фактора роста 1 овцы под промотором гена кератина мыши.
Билет 10
Получение и направление их использования трансгенных коз. ( вроде так)
Нужно для использования их молока для получения медицинских препаратов, поскольку генетические конструкции для экспрессии генов человека в клетках молочной железы этих животных получены и апробированы. В гибридных ДНК используются промоторы гена бетта-лактоглобулина, гена бетта-казеина и гена альфа S1-казеина, а среди клонированных генов человека – ген активатора плазминогена, ген альфа-антитрипсина (при отсутствии этого белка развивается эмфизема, дефицит ингибитора сывороточных протеаз, поражение легких, цирроз печени), ген фактора IX системы свертывания крови (белок нужен для лечения гемофилии В), ген лактоферрина, ген интерлейкина-2. Из трансгенного мелкого рогатого скота упоминают только овец с повышенной скоростью роста шерсти. Этот вариант был получен путем введения кДНК гена инсулиноподобного фактора роста 1 овцы под промотором гена кератина мыши.
Методы введения генетической информации в растения с помощью агробактерий (трансформация изолированных растительных клеток, кокультивация, слияние бактериальных сферопластов и протопластов растительных клеток).
Кокультивация — для получения трансгенных двудольных растений. Для исходного материала нужно иметь штамм агробактерии с векторной конструкцией. Вектор должен содержать последовательность гена, который нужно ввести в геном растения.
В качестве эксплантов для трансформации берут стерильные листовые диски. Экспланты инокулируют жидкой средой. В этой среде находится агробактерия с векторной конструкцией. Происходит заражение клеток раневой поверхности экспланта и после 24-48 ч. культивирования происходит встраивание в растительный геном фрагмента т-ДНК с чужеродным геном. Затем экспланты переносят в стерильную воду и затем подсушивают на поверхности стерильной фильтровальной бумаги (удаление агробактерий). Далее экспланты переносят в жидкую каллусиндуцирующую среду с канамицином и антибиотиком, к которому чувствительны агробактерии (например, ампициллином), и культивируют. После отмывания экспланты переносят на агаризованную каллусиндуцирующую среду с обоими антибиотиками (канамицин + ампициллин) и продолжают культивирование до формирования хорошо выраженных каллусов.
Метод слияния протопластов растительных клеток и сферопластов бактерий. Бактериальные клетки лешенные твердой клеточной стенки называют сферопластами,потому что еще нет способа полного удаления клеточной стенки бактерий. Такие клетки имеют сферическую форму, могут сливаться друг с другом или с протопластами других клеток в растворах веществ, которые способствуют объединению биологических мембран – полиэтиленгликоля, поливиниловый спирта или других. Для реализации такого метода нужно получение сферопластов из бактериальной культуры несущей соответствующую генетическую конструкцию. Штаммы Е.coli можно превратить в сферопласты и их можно использовать для слияния с протопластами растительных клеток.
Трансформация изолированных клеток. Экспременты по введению ДНК в целое растение давали низкую частоту образования трансформантов. Поэтому начали использовать изолированные лишенные клеточной стенки клетки.
Для удаления клеточной стенки растительных клеток используют комплекс ферментов, состоящий из пектинлиаз и целлюлаз. Полученные протопласты трансформируются при обработке их растворами ДНК. Если повысить значение рН растворов до 10 и если в трансформирующих растворах будут находиться дивалентные ионы (Са2+, Mg2+, Zn2+ и Cu2+), то это повысит частоту трансформации.
Лентивирусные векторы: как они стали лучшими векторами для терапии ex vivo
22 октября 2021
Лентивирусные векторы: как они стали лучшими векторами для терапии ex vivo
957
0,0
0
5
Иногда самый безопасный и эффективный вариант генной терапии — не вводить вектор непосредственно пациенту, а наоборот, выделить клетки пациента, произвести с ними генно-инженерные манипуляции и ввести обратно ему же. В таком случае непревзойденный инструмент — лентивирусные векторы. Рисунок в полном размере.
Автор
Редакторы
Возникшая в конце XX века пандемия ВИЧ-инфекции и сопутствующее распространение синдрома приобретенного иммунодефицита (СПИДа) послужили толчком к масштабным исследованиям вызывающего эти патологии вируса. В этих исследованиях раскрыли детали организации генома, репликации и жизненного цикла вируса иммунодефицита человека 1, проложив тем самым путь для создания на его основе лентивирусных векторных систем. Такие системы использовались в создании самой первой одобренной CAR-T-терапии — лечения, подарившего надежду безнадежным больным, и ставшего одним из самых громких прорывов в онкогематологии за последние годы. А помогли тут во многом некоторые уникальные свойства лентивирусов, которые, как выяснилось, могут нести не только смерть, но и жизнь.
Генная и клеточная терапии
Спецпроект о генной и клеточной терапиях, тернистом пути их развития, первых успехах и надеждах, а также о сложностях регулирования, производства и изучения этих новейших методов лечения.
Партнер спецпроекта — Департамент разработки генотерапевтических препаратов одной из крупнейших российских биотехнологических компаний — BIOCAD. BIOCAD заслужил серьезные позиции на мировом фармацевтическом рынке благодаря выпуску лекарственных препаратов на основе антител.
Со времени зарождения генной терапии — начиная с самых первых исследований и клинических испытаний — дорога к ее широкому внедрению была тернистой. Впечатляющие успехи чередовались с досадными провалами, отбросившими на новое лечение порочащую тень, что в какой-то момент резко затормозило развитие области [1].
Вы читаете пятую статью спецпроекта «Генная и клеточная терапии». Ранее мы уже информировали вас, как вообще устроены лекарства будущего [2], а также достаточно подробно останавливались на аденоассоциированных вирусах [3] как важном инструменте «генных терапевтов».
К счастью, сегодня все уже не так драматично: генная терапия стала намного безопаснее — регуляторный надзор за ней усилился, а критерии контроля качества стали строже. И все же, бросая даже беглый взгляд на ландшафт современных видов генного лечения, осознаешь, что путь в клинику для большинства таких лекарств до сих пор крайне непрост. Достаточно сравнить число запускаемых клинических исследований с количеством уже вышедших на рынок терапий — последних несоизмеримо меньше. Причин этому немало: это и вопросы безопасности и эффективности; и сложность разработки и масштабирования производства; и вопросы экономической и практической обоснованности внедрения этих совсем недешевых терапий .
Подробнее о тернистом пути разработки и производства генного лечения читайте в статье: «Генные и клеточные лекарства: сложное регулирование для сложных продуктов» [4]; ну а об экономических перипетиях при внедрении сверхдорогих инновационных лекарств (да и не только их) расскажет статья: «Фармакоэкономика: лечить или экономить?» [5].
Однако генная терапия по-прежнему более чем актуальна, ведь она предлагает регулирование молекулярных процессов на новом уровне, тем самым позволяя подбирать ключи к лечению ранее неприступных болезней.
Существующие ныне генные терапии в основном работают по принципу внедрения нужного генетического материала в определенные клетки пациента, что и позволяет модулировать происходящее в организме с лечебными целями. Поэтому для такого лечения критически важна успешная доставка требуемого гена в целевые клетки и ткани, что чаще всего осуществляют с использованием особых носителей — векторов.
In vivo vs Ex vivo
В предыдущих статьях нашего цикла о генной терапии уже были довольно подробно разобраны имеющиеся ныне решения. Не будем сильно повторяться, но отметим особо важные для дальнейшего повествования моменты.
Обозревая генную терапию, в основном имеет смысл говорить о доставке генов c применением вирусных векторов, — другие способы пока используют гораздо реже.
В рамках такого лечения реализуют два совсем разных подхода: in vivo и ex vivo. Это во многом и определяет выбор подходящих векторов для доставки генов.
Терапия in vivo — это прямой перенос векторов в организм пациента путем внутривенной инфузии раствора, содержащего такие носители. В такой доставке наилучшим образом проявили себя аденоассоциированные векторы, поскольку имеют широкий тропизм — специфичность к различным типам клеток, а также низкий уровень генотоксичности и соответственно бóльшую безопасность [3].
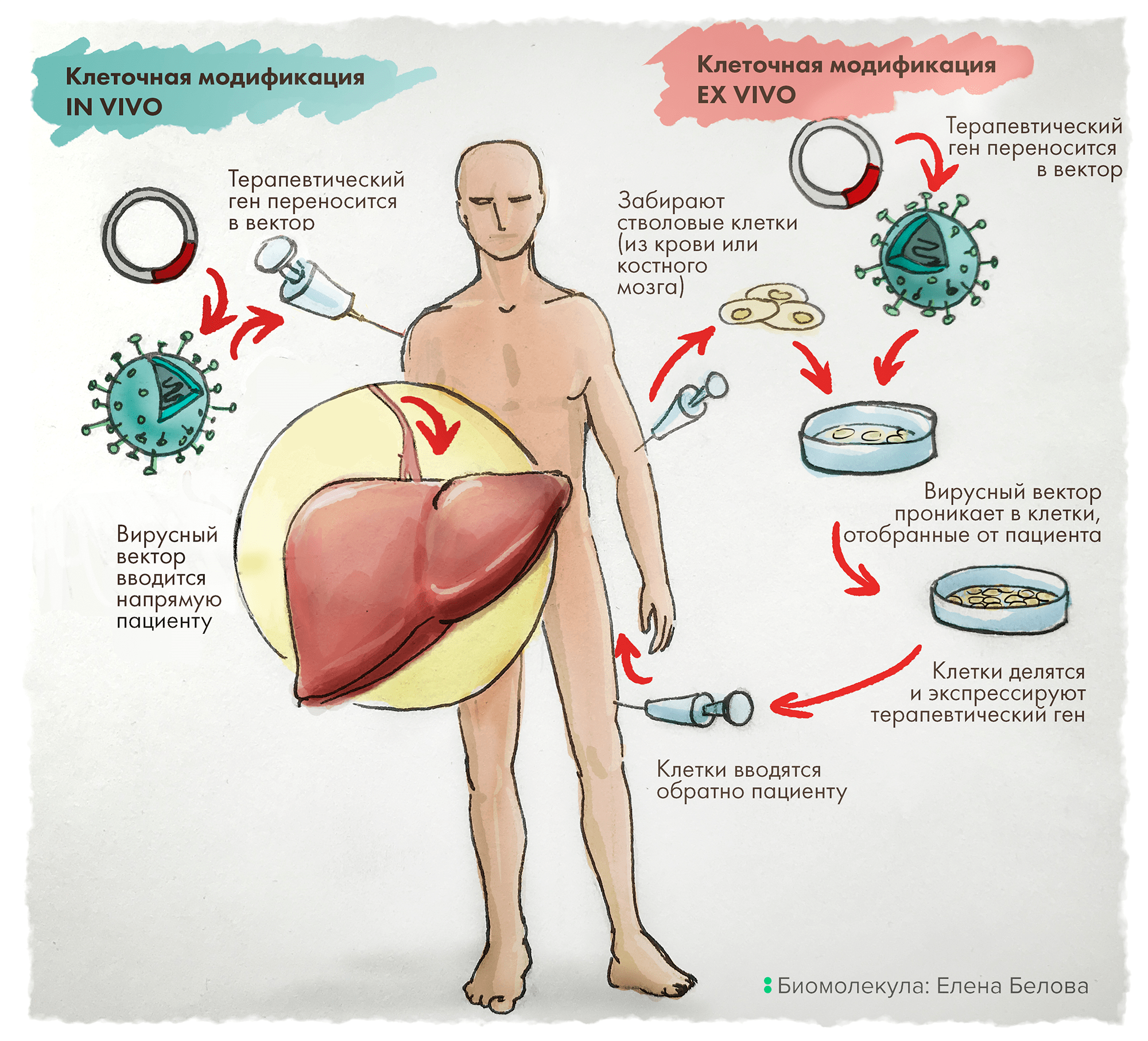
Альтернативно терапевтический ген может быть доставлен ex vivo: для этого клетки сначала отбирают у пациента, генетически модифицируют и культивируют вне организма. И только после этого такие несущие целевые (читай — терапевтические) гены клетки вводят путем инфузии обратно пациенту (рис. 1) [6].
Рисунок 1. In vivo vs Ex vivo. Терапия in vivo заключается в прямой доставке в организм пациента векторов, содержащих терапевтические гены. В отличие от этого, подход ex vivo включает предварительный отбор клеток у пациента, их генетическую модификацию in vitro (это, как правило, внедрение нужного — лечебного — гена), затем отбор эффективно трансдуцированных клеток с последующим размножением и введением такой культуры клеток обратно пациенту.
Для клинических приложений терапии ex vivo изначально были особо интересны γ-ретровирусы и лентивирусы, — ведь они способны перманентно интегрировать свой генетический материал в геном клеток организма-хозяина, что подразумевает передачу генов интереса при делении в ряду клеточных поколений, а значит, низкую элиминацию и долгоиграющую экспрессию.
Однако по сравнению с γ-ретровирусными векторами лентивирусы оказались безопаснее в доклинических исследованиях, а потому сейчас преимущественно используются именно они [7]. Лентивирусные векторы способны еще и трансдуцировать как постмитотические, так и неделящиеся клетки, в то время как для эффективного инфицирования с использованием других ретровирусных платформ требуется, чтобы клетка активно делилась [6]. Дело в том, что среди всех ретровирусов только лентивирусы способны проникать через ядерную мембрану.
Серьезными плюсами лентивирусных векторов являются также относительно низкая стоимость и возможность масштабирования производства. Эта платформа была выбрана для создания одной из наиболее многообещающих ныне генных терапий ex vivo: онколечения с использованием CAR-T-клеток [8], — и используется сегодня при разработке таких препаратов наряду с γ-ретровирусами. К этому мы еще вернемся.
Лентивирусы в природе: что они собой представляют?
Строение
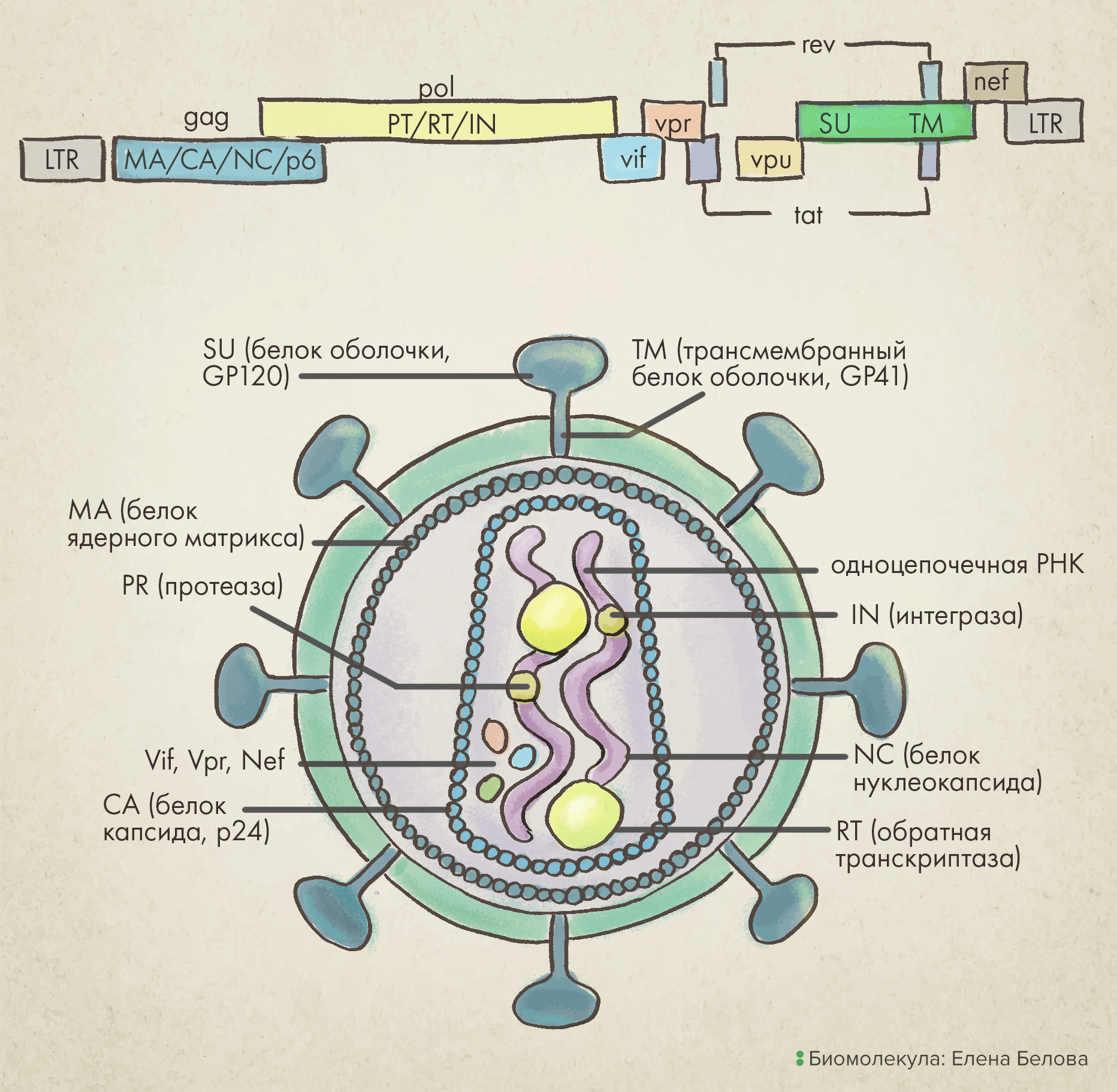
Род лентивирусов из семейства ретровирусов отличается наличием особой оболочки (суперкапсида), внутри которой заключен геном в виде положительной одноцепочечной РНК , которая после попадания вируса в клетку конвертируется в двухцепочечную ДНК. Наиболее известный их представитель — вирус иммунодефицита человека (ВИЧ) [9]. Покрывающий такие вирусы суперкапсид — сферический, слегка плеоморфный, диаметром приблизительно 80–100 нанометров. Длина его генома около 9 килобаз (тысяч пар нуклеотидных оснований), включая три основных структурных гена: gag, pol и env (рис. 2).
Под положительной (+) РНК понимают геномную нуклеиновую кислоту вируса, похожую на мРНК, благодаря чему информация в ходе трансляции — синтеза полипептидной цепи вирусного белка в зараженной клетке — может считываться непосредственно с нее. Такую (+)РНК называют также смысловой. В отличие от этого, антисмысловая (−) РНК комплементарна мРНК, и для перевода в аминокислотную цепочку ее требуется сначала реплицировать в (+)-форму с помощью фермента РНК-репликазы.
Рисунок 2. Лентивирус: геномная карта и структура вириона. Важный для проникновения в клетку ген env обозначен зеленым цветом.
Ген gag первым транскрибируется в незрелую пре-мРНК, которая затем расщепляется на отдельные транскрипты, кодирующие три белка вирусного кора:
Матричный белок (matrix, MA) необходим для сборки вириона и инфекции неделящихся клеток.
Нуклеокапсидный белок (nucleocapsid, NC) накрепко связан с РНК в вирионе — он механически «укрывает» последовательность нуклеотидов, защищая тем самым вирусный геном.
Ген pol кодирует ферменты, необходимые для репликации генетического материала: вирусную протеазу (PRO), обратную транскриптазу (RT) и интегразу (IN).
Наконец, ген env кодирует белок вирусной поверхности: гликопротеин gp160, который в процессе созревания вирусной частицы расщепляется на белок поверхности вириона gp120 (SU) и трансмембранный белок gp41 (TM).
Кроме того, вирусный геном содержит регуляторные гены (tat и rev), которые после инфицирования вирионом экспрессируются первыми и нужны для ускорения продукции мРНК, а также четыре вспомогательных гена (vif, vpr, vpu, и nef) [10].
Жизненный цикл лентивирусов
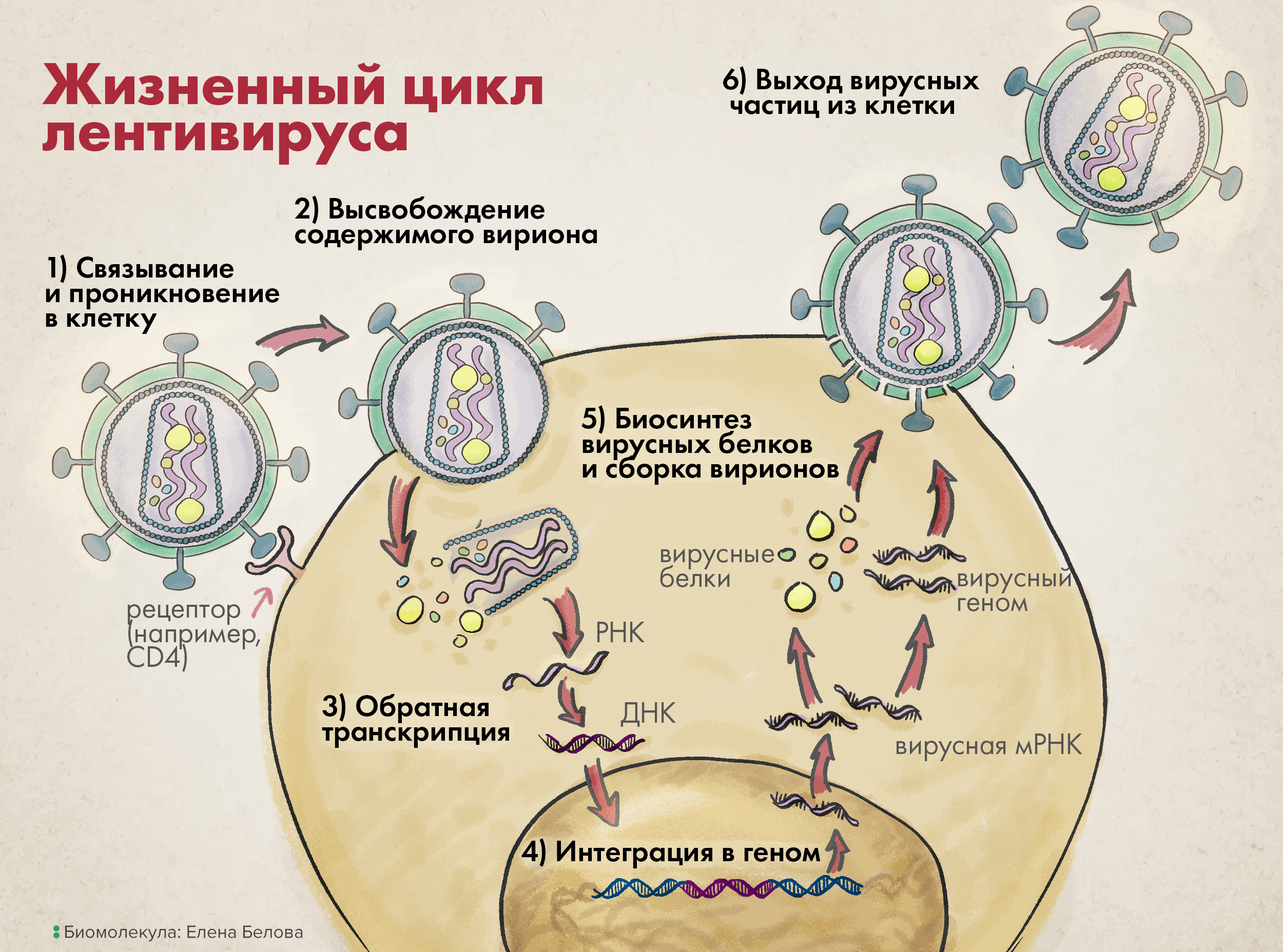
Жизненный цикл лентивируса можно немного обобщить и представить в виде шести основных шагов (рис. 3).
На первой стадии вирион связывается со своей клеткой-мишенью через взаимодействие между белком вирусной оболочки gp120 (SU) и особым рецептором клеточной поверхности.
Существование этих поверхностных и специфичных белков-рецепторов собственно и определяет особую способность вирусов инфицировать строго определенные типы клеток (известную также как тропизм).
Рисунок 3. Жизненный цикл лентивируса.1. Связь с клеточной поверхностью и проникновение в клетку. 2. Высвобождение содержимого вириона во внутриклеточную среду. 3. Обратная транскрипция. 4. Интеграция в геном. 5. Биосинтез вирусных белков и сборка вирионов. 6. Выход вирусных частиц из клетки.
При инициации цикла заражения вирусом ВИЧ, помимо взаимодействия гликопротеина SU с первичным рецептором СD4 на поверхности лимфоцитов Т-хелперов (что и определяет патогенез ВИЧ-инфекции), происходит также его связывание с корецепторами: CXCR4 или CCR5.
Затем вирусные трансмембранные белки (TM) изменяют свою конформацию, чтобы сблизить вирион и клетку, что приводит к слиянию их мембран и проникновению вируса в клетку (этап 1 на рисунке 3).
После входа в клетку матрикс вируса — то есть все его содержимое, кроме слившегося с мембраной клетки суперкапсида, — распределяется в цитоплазме на составляющие элементы: структурные белки капсида, нуклеиновые кислоты вирусного генома, а также ферменты, необходимые для успешной репродукции генетического материала (этап 2 на рисунке 3).
Используя вирусную РНК как шаблон, а также нуклеотиды зараженной клетки, обратная транскриптаза начинает синтезировать комплементарную вирусную ДНК, а точнее — провирусную ДНК (этап 3 на рисунке 3); последняя затем импортируется в клеточное ядро, вставляясь там с использованием вирусной интегразы в геном клетки-мишени (этап 4).
Дальнейшая репродукция генетического материала идет уже как по нотам: в соответствии с центральной догмой молекулярной биологии. Из последовательности провируса в геноме транскрибируются одноцепочечные (+)РНК, затем с них транслируются вирусные белки (этап 5); они упаковывают вирусные нуклеиновые кислоты так, чтобы в одном вирионе таких последовательностей было две (этап 5).
После этого сформированные вирусные частицы, прорывая цитоплазматическую мембрану, выходят из клетки, формируя из ее фрагментов свой суперкапсид (этап 6 на рисунке 3). Ну а такие новые вирионы заражают уже другие клетки, и весь описанный цикл повторяется [10].
Создание лентивирусных векторов
Теперь мы знаем, как структурно устроены лентивирусы, каков их жизненный цикл, но пока еще не разобрались, что собой представляют лентивирусные векторные системы, — как они работают и как их производят. Что же, рассмотрим и этот вопрос.
Безопасность превыше всего
Рекомбинация
Лентивирусы имеют высокую частоту рекомбинации (обмена частями генома между двумя вирусами) и быстро мутируют. Отсюда естественные опасения, что векторы на их основе смогут самопроизвольно воспроизводиться в организме человека и благодаря дальнейшей «перетасовке генов» приводить к появлению суперинфекций. Чтобы снизить подобные риски при разработке векторных систем, основные гены исходных вирусов стали распределять в отдельные плазмиды. При этом полученные векторы (уже неспособные к репликации) — далее еще и последовательно улучшали: на сегодняшний день сменилось уже три поколения таких частиц.
Инсерционный мутагенез
Помимо образования репликационно компетентного вируса, вторым основным риском при применении ретровирусных векторов является онкогенез, возникающий за счет так называемого инсерционного мутагенеза. Критически важно оценивать такие риски, ведь γ-ретровирусы имеют тенденцию встраиваться вблизи генных регуляторных последовательностей, что представляет большую опасность. В частности, в клинических исследованиях с использованием таких частиц фиксировались случаи смерти от развившегося лейкоза [11].
Именно поэтому в поле генной терапии наблюдается постепенное смещение в пользу преимущественного использования лентивирусных векторов, которые намного безопаснее [12]. Они предпочитают транскрипционно активные регионы, но интегрируются в основном в «тело гена» (преимущественно в интроны), избегая энхансеров и промоторов, — что и снижает (и довольно существенно) риски онкогенеза [11], [12].
Поскольку инсерция в транскрипционно активные регионы все же может быть генотоксичной, для усиления биобезопасности разработали так называемые самоинактивирующиеся векторы (self-inactivating, SIN). Они обладают делецией в регионе 3′-LTR вирусного генома: во время обратной транскрипции эта делеция переносится на 5′-LTR провирусной ДНК, что подавляет транскрипционную активность.
Это предотвращает продукцию полноразмерного вируса (после интеграции векторного генома в геном клетки-хозяина), минимизируя тем самым вероятность возникновения репликационно активных вирусных частиц [13]. Более того, такая делеция снижает вероятность, что клеточные кодирующие последовательности, в которые встроился вектор, будут аберрантно экспрессироваться в связи с активностью промотора или через энхансерный эффект. Наконец, дизайн SIN предотвращает потенциальную транскрипционную интерференцию между LTR и промотором, управляющим экспрессией трансгена [13].
Другая повышающая безопасность модификация заключается во включении синтетического инсулятора хроматина в лентивирусные векторы, что снижает взаимодействия между трансгеном и ближайшими к нему генами, также уменьшая возможность аберрантной экспрессии.
Кроме того, правильный подход к дозированию векторов позволяет избегать множественных интеграционных событий в клетке. В идеале требуется подобрать такую дозу, чтобы интеграция происходила в среднем один раз на клетку [12].
В совокупности, все перечисленные меры снижают риск генотоксичности, однако говорить о том, что он сведен на нет, пока еще рано. В нескольких последних исследованиях с использованием лентивирусных векторов были зафиксированы случаи возникновения онкологических заболеваний. В двух из них после проведения расследования испытания продолжили, так как вероятность связи генной терапии и онкозаболеваний посчитали пренебрежимо малой.
Для одного из случаев вопрос о взаимосвязи использования лентивирусных векторов и онкозаболеваний пока остается открытым. Наверное, и мы не будем делать поспешных выводов, ведь с одной стороны, — опасность инсерционного онкогенеза при использовании лентивирусов вполне реальна, а с другой — серьезной статистики по таким случаям нет. Кроме того, в случае очень тяжелых заболеваний, где альтернатива для пациента — постепенное ухудшение состояния и гарантированная смерть через несколько лет, небольшой риск может быть оправдан. Разумеется, ученые при этом не прекращают искать более безопасные способы модификации геномов.
Три поколения векторов
Первые лентивирусные векторные системы содержали три плазмиды: трансферную (transfer), оболочечную (envelope), и упаковочную (packaging).
Упаковочная плазмида содержала бóльшую часть генома исходного вируса: структурные, ферментативные, регуляторные и вспомогательные последовательности, — и не включала гены, кодирующие поверхностные белки вириона. А ведь именно они нужны для связи с клеткой и проникновения в нее, что является ключевым этапом репликативного цикла. Поэтому такие векторные системы (с удаленным геном env) — к репликации уже неспособны, однако уметь проникать в клетку они ведь все же должны — иначе как же они доставят туда целевые гены?
За это отвечает оболочечная плазмида, несущая последовательность вирусного гликопротеина (такого как оболочечный гликопротеин G вируса везикулярного стоматита VSV-G) для помощи векторной частице в связывании с рецепторным белком на поверхности клетки.
Эта технология, основанная на общем свойстве большинства ретровирусов, — возможности формировать так называемые псевдотипы (частицы, в оболочку которых включается поверхностный белок какого-то другого вируса) — позволила расширить диапазон поражаемых ими клеток. Псевдотипированные белком VSV-G векторы обладают уникальными преимуществами: большей стабильностью, а также способностью связываться с рецепторами LDL (липопротеинов низкой плотности), присутствующим на поверхности множества клеток, — что и определяет широкий тропизм этих частиц [10]. Существуют и другие варианты псеводтипирования векторов, расширяющие их тропизм (табл. 1).
Таблица 1. Различные псевдотипы лентивирусных векторов, гликопротеины их вирусных оболочек, и свойственный таким псевдотипам клеточный тропизм.Источник: [14].
Лентивирусные псевдотипы
Исходный вирус (у которого «позаимствован» поверхностный гликопротеин вирусной оболочки)
Клеточный рецептор, с которым связывается псевдотип
Клеточный тропизм
Эффективность
VSV-G
Вирус везикулярного стоматита
LDL-R
Широкий тропизм в не первичных (подвергнутых хотя бы одному пассивированию) культурах клеток
Высокая
BaEV
Эндогенный ретровирус павиана
ASCT-1 ASCT-2
CD34+ клетки
30%
Наивные Т-клетки
До 80%
Наивные В-клетки
40%
В-клетки памяти
20%
Натуральные киллеры
40%
Ранние тимоциты
До 80%
RD114
Эндогенный ретровирус кошек
ASCT-2
Наивные Т-клетки
До 60%
Наивные В-клетки
До 30%
H/F
Вирус кори
SLAM CD46
CD34+ клетки
──
Покоящиеся Т-клетки памяти
──
Наивные Т-клетки
До 50%
Покоящиеся В-клетки
Покоящиеся гемопоэтические стволовые клетки
До 70%
Дендритные клетки
G/F
Вирус Нипах
EphinB2/B3
Перициты
20–40%
Клетки эндотелия опухоли
COCV
Везиколовирус Кокал
LDL-R
Стимулированные клетки CD34+
До 80%
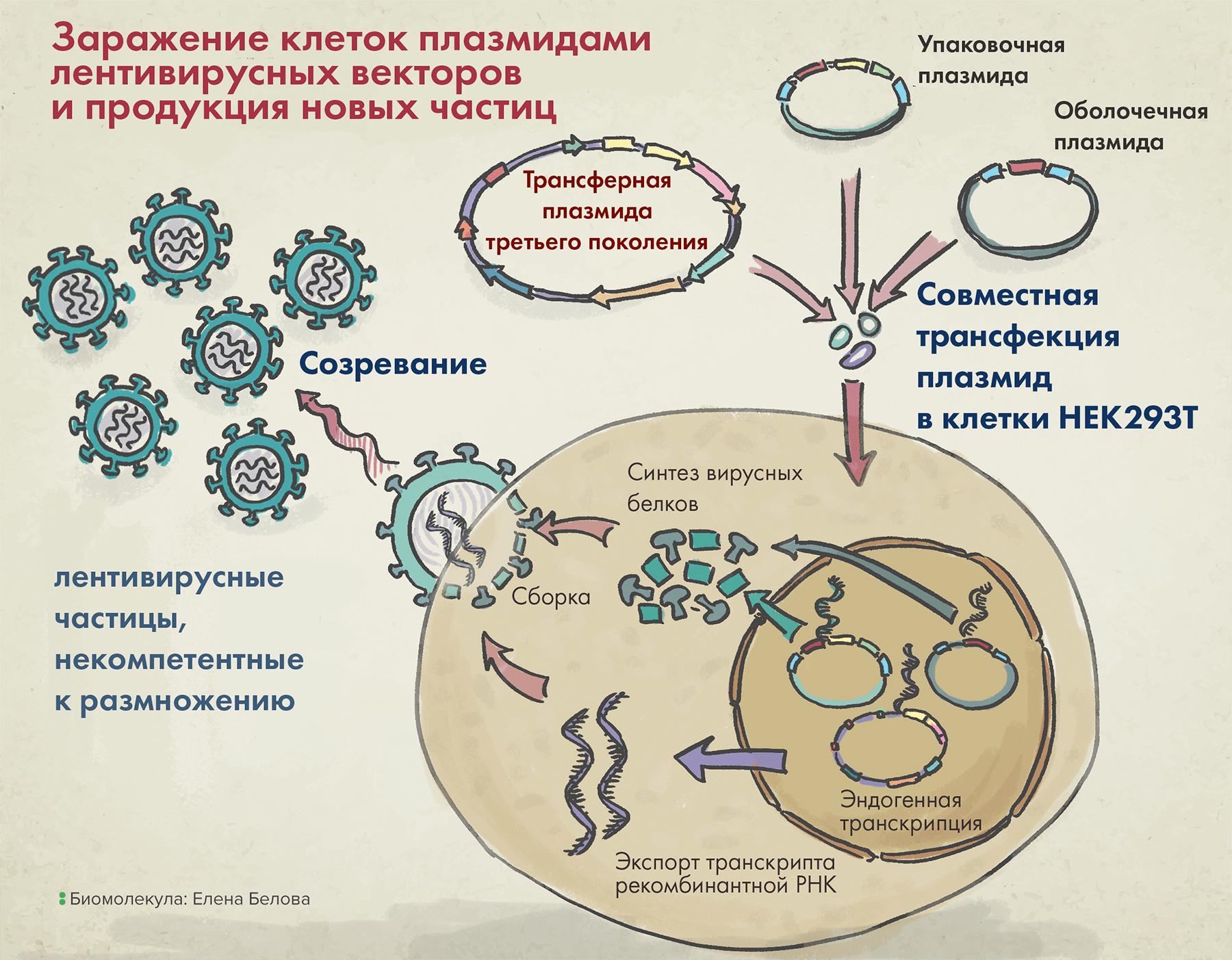
Наконец, последний компонент векторной системы первого поколения — трансферная плазмида — несет целевой трансген, а также так называемую последовательность пси-петли (ψ-петли), — и все это в обрамлении длинных терминальных повторов (long terminal repeats, LTRs). Последние два элемента необходимы для упаковки вирусной РНК в вирион (рис. 4).
Рисунок 4. Упаковка лентивирусных векторов в клетке (на примере третьего поколения таких систем).
После заражения клеток тремя вышеназванными плазмидами в них происходит сборка вирусных частиц, а затем уже их выход наружу. Высвобождаемые векторы содержат VSV-G (он помогает им впоследствии проникать в клетки), но внутри них нет гена, кодирующего этот белок. Так происходит потому, что в кодирующей VSV-G плазмиде отсутствуют генетические последовательности, необходимые для сборки белковой вирусной оболочки, а значит, все ее содержимое запаковаться в вирион уже не сможет.
Поэтому такие частицы умеют доставлять гены в клетку, но не способны размножаться. Чтобы они вдруг снова научились этому, необходима рекомбинация между нашими векторами и, например, природными лентивирусами. Гипотетически это возможно, хоть и маловероятно, — ведь для восстановления репликативно компетентного генома должно многое совпасть: одновременное заражение клеток и вектором, и вирусом, а также прицельный обмен между ними генетическим материалом, — что в силу случайного характера рекомбинации вряд ли случится.
Тем не менее чтобы обезопаситься еще больше, исследователи стали менять исходный геном лентивируса еще сильнее: разбивать его на большее число плазмид, удалять еще больше нецелевых генов, а также модифицировать все необходимые в векторе нативные последовательности. Такие стратегии легли в основу создания лентивирусных векторов второго и третьего поколений (табл. 2).
Таблица 2. Второе и третье поколения лентивирусных векторов.Источник: Merck.
Характеристика
Второе поколение
Третье поколение
Количество плазмид
3
4
Наличие делеции в области U3 в 3′ LTR — для создания самоинактивирующегося вектора (Self-INactivation, SIN)
Нет
Да
5′ LTR
Дикого типа (5′ LTR)
Химерные (RSV)
Количество упаковочных плазмид
1
2
Вспомогательные гены: vif, vpr, vpu, nef
Отсутствуют
Отсутствуют
tat и rev гены
В упаковочной плазмиде
Ген tat удален, rev в отдельной упаковочной плазмиде
gag и pol гены
В упаковочной плазмиде, вместе с tat и rev
Вместе в отдельной плазмиде
Псевдотипирование
Псевдотипированы (VSV-G)
Псевдотипированы (VSV-G)
Количество рекомбинационных событий, необходимых для генерации репликационно компетентных вирусов
3 рекомбинации
4 рекомбинации
Производство
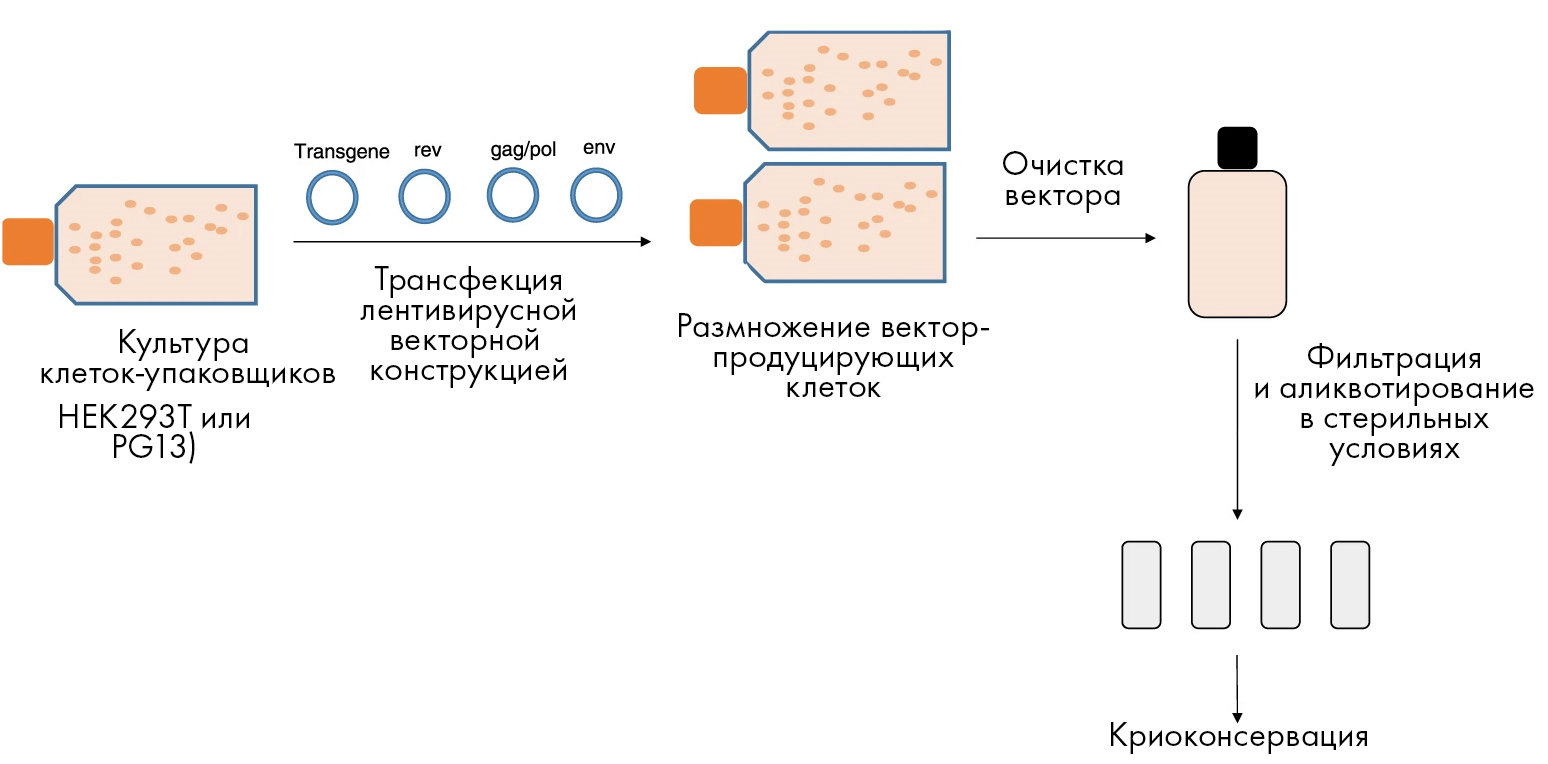
Создание эффективных векторных систем — это лишь первый этап к достаточной продукции нужного материала. Не менее важна разработка технологии производства и ее последующее масштабирование на большие объемы, — чтобы можно было получать требуемые частицы в нужных количествах. При этом в современном мире продукция вирусных частиц для лечебного применения должна идти по Правилам надлежащей производственной практики (известной также как GMP). На рисунке 5 даны основные стадии производства лентивирусных векторов.
Рисунок 5. Основные этапы продукции вирусных векторов на современном производстве, работающем в соответствии с принципами GMP. Все начинается с культуры специальных «клеток-упаковщиков» (таких как HEK293T или PG13). Эти особые упаковочные линии культивируют в течение нескольких дней, заражают плазмидами третьего поколения лентивирусного вектора, и затем уже такие вектор-продуцирующие клетки снова размножают в культуре. После этого векторы выделяют и очищают от остатков клеток (т.н. дебриса: различных клеточных органелл, плазмалеммы и т.д.), а также других загрязняющих примесей. После очистки содержащий векторы полупродукт фильтруют через мембрану с определенным размером пор, не пропускающюю микроорганизмы (что позволяет достичь стерильности). Далее полученный сток асептично распределяют по отдельным аликвотам и криоконсервируют. Сообщалось, что такие замороженные при −80 °С векторы могут сохранять свою стабильность до 9 лет после криоконсервации. Стоит также отметить, что в течение всего описанного процесса очень важен контроль качества, ведь, например, не до конца удаленный из стоков вектора клеточный дебрис может вызывать воспалительные реакции, что было показано в исследованиях как in vitro, так и in vivo [15].
Альтернатива: стабильные упаковщики
Еще один способ получать лентивирусные векторы, помимо временной трансфекции плазмидами, — создание стабильных линий клеток-упаковщиков. Для этого геном клеток (чаще всего это клетки почки человеческого эмбриона HEK293T) модифицируют так, чтобы они смогли стабильно экспрессировать бóльшую часть вирусного генетического материала.
Такие клетки-упаковщики, как правило, экспрессируют гены gag, pol и rev, а зачастую также ген капсида VSV-G. Далее для использования в качестве продуцентов векторов их нужно заразить плазмидой, несущей целевой ген.
Пока что такие стабильные линии используют реже чем «стандартные» клетки-продуценты, ─ но перспективы расширения их применения весьма заманчивы. Они могут осуществлять долгоиграющую экспрессию, что крайне желательно, ─ так как позволит упростить производство, снизив тем самым и его стоимость [16].
Применение лентивирусных векторов в клинике
С момента первых применений лентивирусные векторы стали одним из основных инструментов арсенала генной и клеточной терапии с заметными успехами в клинических исследованиях [17]. Одна из причин — они изначально обладали большим потенциалом благодаря способности эффективно трансдуцировать CD34 + стволовые клетки. Из последних, как известно, в процессе гемопоэза появляются все остальные типы кровяных клеток [18], а потому их генетическая модификация была крайне привлекательна. Ведь после дифференциации эти измененные клетки теоретически смогут остановить (или хотя бы притормозить) патогенез самых разных связанных с ними болезней.
Ученым и медикам оставалось лишь проверить эту теорию практикой, и вскоре перенос генов с использованием лентивирусного вектора в CD34+ стволовые клетки был осуществлен для лечения нескольких генетических заболеваний. Среди них: бета-талассемия, серповидноклеточная анемия, Х-сцепленная адренолейкодистрофия, метахроматическая лейкодистрофия, синдром Вискотта—Олдрича и др. При этом о наличии каких-либо связанных с вектором серьезных нежелательных явлениях в этих исследованиях заявлено не было (о первом тревожном сигнале, связанным с онкогенезом, рассказано выше) [15].
Ранние успехи подстегнули развитие такого лечения, и, согласно регистру клинических испытаний ClinicalTrials.gov, функциональность лентивирусного вектора сейчас уже тестируют более чем в 4000 исследований. При этом известно, что и количество подобных испытаний непрерывно и экспоненциально растет, например: в 2017 г. их было около 200, в 2020-м — более 600 [19], ну а сегодня, как видим, — уже на порядок больше. Из такого обширного поля настоящими прорывами можно назвать уже одобренное лечение, — а это пока что два генных продукта:
Zynteglo (Bluebird bio, Массачусетс, США) для лечения бета-талассемии.
Kymriah (Novartis, Базель, Швейцария) для лечения онкогематологических болезней [19].
Они оба представляют собой лечение ex vivo аутологичными генномодифицированными клетками крови пациентов, однако Novartis использует еще и принципиально новый (хоть и разрабатываемый немало лет) подход — внедрение генов для изменения поверхностного рецептора T-лимфоцитов. После модификации такой рецептор способен распознавать специфичные маркеры на опухолевых клетках, — ну а уже измененные T-клетки пациентов уничтожают злокачественные бласты .
Заключение и перспективы
Одно из направлений улучшения генной терапии — разработка новых векторов-носителей. В частности, представляют интерес не интегрирующиеся лентивирусные векторы (non-integrating lentiviral vectors, NILVs) — они привлекательны в плане снижения вероятности инсерционного мутагенеза. В них выключен фермент вирусная интеграза (как мы помним, отвечающая за внедрение провирусной последовательности в геном клетки) — и они точно так же умеют инфицировать как делящиеся, так и неделящиеся клетки. При этом вирусный геном остается в клетке в виде эписомы, а не интегрируется в геномную ДНК [15].
NILVs стабильно экспрессируют трансгены из эписомальной ДНК в неделящихся клетках, и временно — если клетки-мишени размножаются. Было показано, что такие векторы имеют широкий тропизм ко множеству различных типов клеток и тканей, что вкупе с повышенной безопасностью делает их идеальными кандидатами для использования в вакцинации и иммунотерапии.
Это также открывает перспективы использования NILVs как инструмента для доставки компонентов CRISPR/Cas9 — революционной технологии, которая ныне широко используется в исследованиях генного редактирования. Она включает так называемую направляющую РНК, помогающую распознавать нужные участки генома в клетке для связи с ними нуклеазы Cas9. Последняя начинает в этом месте расщеплять спираль ДНК, создавая двухцепочечные разрывы, которые затем могут быть репарированы определенным образом — с удалением (делецией) каких-либо «поломанных» генов, или внедрением в этом месте каких-то новых кодирующих последовательностей.
Сегодня существуют лентивирусные системы, умеющие доставлять в клетку как направляющие РНК, так и нуклеазу Cas9 (точнее кодирующие их последовательности — для последующей экспрессии).
Лентивирусные векторы также часто используют в исследовательских целях для изменения экспрессии генов посредством использования короткой шпилечной РНК или антисмысловой РНК [23], — что является частью уже известного механизма РНК-интерференции (об этом интересном феномене «Биомолекула» писала уже не раз).
Конечно, пока что большинство из таких перспективных подходов все еще находится на ранних стадиях разработки, и необходимы дальнейшие исследования, чтобы определить, насколько лентивирусные векторы по настоящему «жизнеспособны» — как инструмент для терапевтической доставки компонентов редактирования/регуляции экспрессии тех или иных генов [15].